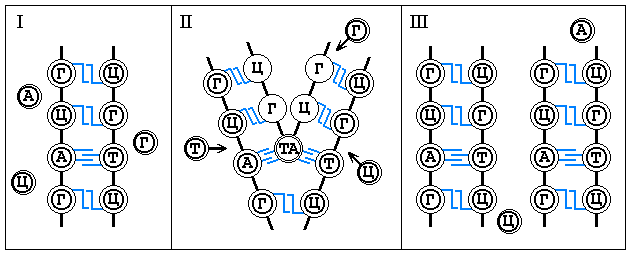
Blagodarya principu komplementarnosti, lezhashchemu v osnove
postroeniya dvojnoj spirali DNK, molekula DNK sposobna k udvoeniyu. Vo vremya etogo
processa dve cepi raz®edinyayutsya, obrazuya pri etom dve dvojnye cepi fn. yacheek, u kotoryh
lish' odin ryad zapolnen fshch. edinicami, a drugoj stanovitsya svobodnym. Na sleduyushchem etape
neassociirovannye nukleotidy iz okolosistemnoj sredy zapolnyayut sootvetstvuyushchie im
svobodnye fn. yachejki v obeih spiralyah. V rezul'tate reduplikacii vmesto odnoj molekuly
DNK voznikaet dve takogo zhe tochno nukleotidnogo sostava, kak i pervonachal'naya. Odna cep'
v kazhdoj vnov' obrazovavshejsya molekule DNK ostaetsya ot pervonachal'noj molekuly, drugaya
sinteziruetsya vnov'. Takim putem vmeste so strukturoj proishodit peredacha fn. svojstv
DNK ot materinskoj kletki k dochernej.
Graficheski eto vyglyadit sleduyushchim obrazom:
Molekuly RNK takzhe yavlyayutsya polimerami, kak i DNK,
no v otlichie ot nih imeyut odnu spiral' fn. yacheek, a ne dve. RNK vypolnyaet v kletke
ryad funkcij, v tom chisle
1) transportnuyu (transportiruyut
aminokislotnye bloki k mestu sinteza belkov);
2) informacionnuyu (perenosyat
informaciyu o strukture belka);
3) ribosomnuyu.
Eshche odnim ochen' vazhnym nukleotidom v strukture zhivoj kletki
yavlyaetsya adenozintrifosfornaya kislota - ATF, soderzhanie kotoroj v kletkah kolebletsya
ot 0,04 do 0,2 - 0,5%. Ego osobennost' sostoit v tom, chto pri otshcheplenii odnoj molekuly
fosfornoj kisloty ATF perehodit v ADF (adenozindifosfornuyu kislotu) s vydeleniem 40 kdzh
energii s 1 gr-molekuly.
Vse ukazannye vyshe organicheskie veshchestva yavlyayutsya slozhnymi po
svoej strukture i sistemnoj organizacii obrazovaniyami, no i oni, v svoyu ochered', vhodyat
v kachestve fshch. edinic v fn. podsistemy integrirovannoj sistemy kletki. K chislu osnovnyh
podsistem kletki otosyatsya sleduyushchie:
Naruzhnaya membrana kletki. Reguliruet postuplenie
ionov i molekul v strukturu kletki i vyhod ih iz nee v okolosistemnuyu sredu. Takoj obmen
molekulami i ionami, to est' razlichnymi fshch. edinicami, mezhdu sistemoj kletki i vneshnej
sredoj proishodit postoyanno. Razlichayut fagocitoz - pogloshchenie membranoj krupnyh chastichek
veshchestva i pinocitoz - pogloshchenie vody i vodnogo rastvora. CHerez naruzhnuyu membranu iz
kletki vyvodyatsya produkty ee zhiznedeyatel'nosti, to est' otfunkcionirovavshie v podsistemah
kletki fshch. edinicy.
Citoplazma. Vnutrennyaya poluzhidkaya sreda kletki,
v sistemnom ob®eme kotoroj razvernuta vnutrennyaya struktura kletki, to est' ee yadro, vse
organoidy (ili organelly), vklyucheniya i vakuoli. Citoplazma sostoit iz vody s rastvorennymi
solyami i raznoobraznymi organicheskimi veshchestvami, sredi kotoryh preobladayut belki.
Struktura citoplazmy sostoit iz fn. yacheek, ne svyazannyh zhestko i svobodno peremeshchayushchihsya
vo vsem ee ob®eme. Zapolnyayushchie ih fshch. edinicy, kogda eto neobhodimo, perehodyat iz nih v
fn. yachejki organoidov. Poetomu osnovnymi funkciyami citoplazmy yavlyayutsya akkumulyativnaya i
transportnaya.
|ndoplazmaticheskaya set'. Organoid kletki,
predstavlyayushchij soboj slozhnuyu sistemu kanalov i polostej, pronizyvayushchih vsyu citoplazmu
kletki. Na membranah gladkoj endoplazmaticheskoj seti proishodit sintez zhirov i uglevodov,
kotorye nakaplivayutsya v akkumulyativnyh fn. yachejkah ee kanalov i polostej, a zatem
transportiruyutsya k razlichnym organoidam kletki, gde oni v kachestve fshch. edinic zanimayut
sootvetstvuyushchie fn. yachejki ih struktur. Na membranah kanalov i polostej raspolagaetsya
takzhe mnozhestvo melkih okruglyh telec-ribosom.
Kazhdaya ribosoma sostoit iz dvuh melkih chastic, v sostav kotoryh
vhodyat belki i RNK. V kazhdoj kletke soderzhitsya po neskol'ku tysyach ribosom. Na ribosomah
sinteziruyutsya vse belki, vhodyashchie v sostav dannoj kletki, putem sborki belkovyh molekul
iz aminokislot, imeyushchihsya v citoplazme. Sintez belkov - eto slozhnyj process zapolneniya
aminokislotnymi blokami sootvetstvuyushchih fn. yacheek ih struktur, kotoryj osushchestvlyaetsya
odnovremenno gruppoj iz neskol'kih desyatkov ribosom, ili poliribosomoj. Sintezirovannye
belki snachala nakaplivayutsya v kanalah i polostyah granulyarnoj endoplazmaticheskoj seti, a
zatem transportiruyutsya k tem podsistemam kletki, gde raspolozheny prednaznachennye dlya nih
fn. yachejki. |ndoplazmaticheskaya set' i poliribosomy predstavlyayut soboj edinyj mehanizm
biosinteza, akkumulirovaniya i transportirovki belkov.
Mitohondrii. Organoid, osnovnaya funkciya
kotorogo sostoit v sinteze ATF, predstavlyayushchej universal'nyj istochnik energii, neobhodimoj
dlya osushchestvleniya postyanno protekayushchih vnutri kletki himicheskih processov. Kolichestvo
mitohondrij v kletke kolebletsya ot neskol'kih do soten tysyach. Vnutri mitohondrij nahodyatsya
ribosomy i nukleinovye kisloty, a takzhe bol'shoe kolichestvo raznoobraznyh fermentov.
Sintezirovannaya ATF zapolnyaet transportnye fn. yachejki citoplazmy i napravlyaetsya k yadru
i organoidam kletki.
Plastidy. Organoidy rastitel'nyh kletok.
Byvayut razlichnyh tipov. S pomoshch'yu odnogo iz nih - hloroplastov, blagodarya vhodyashchemu
v ih sostav pigmentu (hrolofillu), kletki rastenij sposobny, ispol'zuya svetovuyu energiyu
Solnca, sintezirovat' iz neorganicheskih veshchestv organicheskie (uglevody). |tot process,
kak izvestno, nosit nazvanie fotosinteza.
Kompleks Gol'dzhi. Organoid vseh rastitel'nyh
i zhivotnyh kletok, v kotorom proishodit nakoplenie belkov, zhirov i uglevodov s posleduyushchej
ih transportirovkoj v sootvetstvuyushchie fn. yachejki kak vnutri kletki, tak i vne ee.
Lizosomy. Organoid, imeyushchijsya vo vseh kletkah
i sostoyashchij iz kompleksa fermentov, sposobnyh rasshcheplyat' belki, zhiry i uglevody. V etom
zaklyuchaetsya osnovnaya funkciya lizosom. V kazhdoj kletke sosredotocheny desyatki lizosom,
uchastvuyushchih v rasshcheplenii uzhe otfunkcionirovavshih ili akkumulyativnyh sistemnyh
obrazovanij, a takzhe teh, kotorye popadayut v kletku izvne putem fagocitoza i pinocitoza.
V rezul'tate rasshchepleniya fshch. edinicy pokidayut fn. yachejki rasshcheplyaemyh struktur, sobirayutsya
v fn. yachejkah akkumulyativnyh sistem dannoj kletki, a zatem transportiruyutsya v fn. yachejki
ee novyh sistemnyh obrazovanij. Rasshcheplennye s pomoshch'yu lizosom otfunkcionirovavshie
struktury kletki udalyayutsya za ee predely. Obrazovanie novyh lizosom proishodit v kletke
postoyanno. Fermenty, funkcioniruyushchie v lizosomah, kak i vsyakie drugie belki, sinteziruyutsya
na ribosomah citoplazmy. Zatem eti fermenty postupayut po kanalam endoplazmaticheskoj seti
k kompleksu Gol'dzhi, v polostyah i trubochkah kotorogo formiruyutsya fn. yachejki struktur
lizosom. Sformirovavshis', lizosomy otdelyayutsya ot koncov trubochek i postupayut v citoplazmu.
Kletochnyj centr. Organoid, raspolozhennyj v
odnom iz uchastkov uplotnennoj citoplazmy. V nego vhodyat dve centrioli, igrayushchie vazhnuyu
rol' pri delenii kletki.
Struktura kletok soderzhit i drugie organoidy: zhgutiki,
resnichki i t.p., a takzhe kletochnye vklyucheniya (uglevody, zhiry, belki).
Vmeste s tem, kletki, buduchi sami po sebe ochen' slozhnymi
sistemnymi obrazovaniyami, v svoyu ochered' yavlyayutsya fshch. edinicami, zapolnyayushchimi fn.
yachejki gipersistem posleduyushchih urovnej organizacii Materii. Vsledstvie etogo v
sistemnoj organizacii kletok predusmotren mehanizm, pozvolyayushchij za sravnitel'no
korotkij period vremeni sozdavat' analogichnye im sistemnye formirovaniya. V rezul'tate
kletochnyj cikl vklyuchaet dva perioda:
1) Delenie (mitoz), v processe kotorogo
obrazuyutsya dve dochernie kletki;
2) Period mezhdu dvumya deleniyami - interfaza -
sobstvenno vremya funkcionirovaniya kletki.
Bol'shuyu rol' v delenii kletki igraet ee yadro, imeyushcheesya
v kazhdoj kletke i predstavlyayushchee soboj slozhnuyu fn. podsistemu. YAdro imeet yadernuyu
obolochku, cherez kotoruyu v nego i iz nego postupayut belki, uglevody, zhiry, nukleinovye
kisloty, voda i raznoobraznye iony. Popav vnutr' yadra, oni zapolyayut fn. yachejki yadernogo
soka, a takzhe yadryshek i hromatina. V yadryshkah proishodit sintez RNK, sami zhe oni
formiruyutsya tol'ko v interfaze. Hromatin predstavlyaet soboj odnorodnoe veshchestvo,
sluzhashchee akkumulyativnoj podsistemoj, s pomoshch'yu kotoroj osushchestvlyaetsya formirovanie
hromosom pri delenii yader.
Hromosomy yavlyayutsya osnovnym mehanizmom kletki, gde
sobiraetsya, hranitsya i vydaetsya tak nazyvaemaya nasledstvennaya informaciya, vklyuchayushchaya
v sebya himicheskuyu zapis' posledovatel'nosti fn. yacheek v strukturah belkov dannoj kletki.
Ukazannaya informaciya hranitsya v nahodyashchihsya v hromosomah molekulah DNK. Takim obrazom,
molekuly DNK predstavlyayut soboj himicheskuyu zapis' struktur vsego raznoobraziya belkov.
Na dlinnoj niti molekuly DNK odna za drugoj sleduet zapis' informacii o
posledovatel'nosti fn. yacheek struktur raznyh belkov. Otrezok DNK, soderzhashchij informaciyu
o strukture odnogo belka, prinyato nazyvat' genom. Molekula DNK predstavlyaet soboj
sobranie neskol'kih sot ili tysyach genov. Diametr hromosom nevelik i sostavlyaet v srednem
140  , dlina zhe
ih, povtoryaya dlinu molekul DNK, mozhet byt' svyshe 1 mm. V seredine perioda interfazy
proishodit sintez DNK, v rezul'tate kotorogo hromosoma udvaivaetsya.
, dlina zhe
ih, povtoryaya dlinu molekul DNK, mozhet byt' svyshe 1 mm. V seredine perioda interfazy
proishodit sintez DNK, v rezul'tate kotorogo hromosoma udvaivaetsya.
Vazhnejshaya funkciya hromosom - byt' hranitel'nicej zapisej
struktur i, sootvetstvenno, algoritmicheskih sposobnostej fn. podsistem kletki s pomoshch'yu
mehanizma obrazovaniya belkovyh fshch. edinic. S techeniem vremeni, po mere prirashcheniya funkcij
togo ili inogo vida organicheskih sistem, izmenyaetsya i sovershenstvuetsya zapis' v hromosomah,
sleduya trebovaniyam zakonov fn. razvitiya Materii. V pryamoj zavisimosti ot molekulyarnoj
zapisi DNK hromosom cherez mehanizm sintezirovaniya belkovyh molekul proishodyat vse
processy zhiznedeyatel'nosti kletok. CHislo hromosom postoyanno dlya kazhdogo vida zhivotnyh
i rastenij, to est' v kazhdoj kletke lyubogo organizma, prinadlezhashchego k odnomu vidu,
soderzhitsya sovershenno opredelennoe chislo hromosom (rozh' - 14, chelovek - 46, kurica -
78 i t.d.). Hromosomnyj nabor, soderzhashchijsya v yadre odnoj kletki, imeet vsegda parnye
hromosomy. Tak, 46 hromosom cheloveka obrazuyut 23 pary, v kazhdoj iz kotoryh ob®edineny
dve odinakovye hromosomy. Hromosomy raznyh par otlichayutsya drug ot druga po forme i mestu
raspolozheniya. V rezul'tate mitoza poluchayutsya dve dochernie kletki, po stroeniyu polnost'yu
shodnye s materinskoj. Kazhdaya iz nih imeet tochno takie zhe hromosomy i takoe zhe ih chislo,
kak i materinskaya kletka. Takim putem obespechivaetsya polnaya peredacha vsej nasledstvennoj
informacii kazhdomu iz dochernih yader. YAdro i vse organoidy citoplazmy kletki
vzaimodejstvuyut kak edinaya sistema.
Vse kletki imeyut shodnyj tip stroeniya: yadro, mitohondrii,
kompleks Gol'dzhi, endoplazmaticheskuyu set', ribosomy i drugie organoidy. Odnako, prezhde
chem stat' stol' sovershennoj sistemoj, kakoj ona yavlyaetsya v nashi dni, kletka proshla
dlitel'nyj put' evolyucii, otmechennyj sootvetstvuyushchimi otrezkami na ordinatah t i
ft tenzora Razvitiya Materii. Vnachale ona byla chast'yu neizvestnyh nam
nekletochnyh organizmov, zatem nesovershennyh odno- i mnogokletochnyh, vklyuchaya bakterii
i sinezelenye vodorosli, i v konechnom itoge dostigla sovershenstva slozhnogo kletochnogo
mehanizma, harakternogo dlya predstavitelej sovremennogo nam rastitel'nogo i zhivotnogo
mira. Vsledstvie dvizheniya Materii po ordinate kachestva v processe evolyucii kletki
obrazovalos' ogromnoe raznoobrazie ee tipov, kazhdyj iz kotoryh byl nadelen strogo
opredelennymi fn. svojstvami i sootvetstvuet opredelennoj tochke na dannoj ordinate.
Vmeste s tem, s opredelennogo momenta etot process stal
protekat' odnovremenno s nachalom razvitiya fn. sistem bolee vysokogo organizacionnogo
urovnya, fn. yachejki kotoryh kletki stali zapolnyat' v kachestve fshch. edinic. V itoge kletka
prevratilas' v slozhnoe sistemnoe obrazovanie, dlya podderzhaniya fn. svojstv kotorogo vnutri
i vne ee postoyanno protekayut slozhnye himicheskie processy. Postoyanstvo processov svyazano s
tem, chto vremya funkcionirovaniya fshch. edinic po mere uvelicheniya ih molekulyarnogo vesa vse
bolee ne sovpadaet so vremenem sushchestvovaniya fn. yacheek struktur, kotorye oni zapolnyayut,
tak kak pri ogranichennom prostranstve peremeshcheniya fshch. edinic vremya ih sushchestvovaniya
nahoditsya v pryamoj zavisimosti ot ih fshch. massy. Krome togo, postoyanstvo processov vyzvano
tem, chto bol'shinstvo protekayushchih v kletke himicheskih reakcij imeet neobratimyj harakter.
Dlya vseh etih reakcij svojstvenny velichajshaya organizovannost' i uporyadochennost': kazhdaya
reakciya protekaet v strogo opredelennom meste v strogo opredelennoe vremya v strogo
opredelennoj posledovatel'nosti. Molekuly fermentov raspolozheny na membranah mitohondrij
i endoplazmaticheskoj seti v tom poryadke, v kotorom idut reakcii.
V kletke imeyutsya okolo tysyachi fermentov, s pomoshch'yu kotoryh
protekayut dva tipa reakcij: sinteza i rasshchepleniya. Osnovnym (sozidayushchim)
tipom reakcij sleduet schitat' reakcii sinteza, v processe kotoryh obrazuyutsya slozhnye
molekulyarnye soedineniya, zapolnyayushchie v kachestve fshch. edinic fn. yachejki podsistemnyh
struktur kletki. Tak, na zamenu kazhdoj otfunkcionirovavshej molekuly belka, pokinuvshej
tu ili inuyu fn. yachejku, na osvobodivsheesya mesto postupaet novaya molekula belka, po
strukture i himicheskomu sostavu, a sledovatel'no i svoim fn. svojstvam, polnost'yu
adekvatnaya predydushchej fshch. edinice. |to oznachaet, chto vnov' sintezirovannaya fshch. edinica
sposobna (ili dolzhna byt' sposobna) prinyat' polnocennoe uchastie v lyubom iz harakternyh
dlya dannoj fn. yachejki algoritmov.
Sintezirovanie fshch. edinic osushchestvlyaetsya s pomoshch'yu
funkcionirovaniya special'nyh podsistem kletki na osnove kodirovannoj genozapisi DNK.
Imeyushchie pri etom mesto fluktuacionnye otkloneniya v sluchae ih polozhitel'nogo effekta
obratnoj svyaz'yu fiksiruyutsya v genozapisi i sluzhat celyam dal'nejshego sovershenstvovaniya
dannoj sistemnoj struktury. Pri otricatel'nom effekte ot vnov' sintezirovannoj fshch.
edinicy narushaetsya vypolnenie chasti fn. algoritmov i, v sluchae, esli sistema ne v
sostoyanii eto eliminirovat', nenadlezhashchee funkcionirovanie sootvetstvuyushchej podsistemy
v konechnom itoge mozhet privesti k razrusheniyu struktury dannoj kletki v celom. Takim
putem organizacionnoe ustrojstvo kletki pozvolyaet ej podderzhivat' postoyannoe nalichie
sootvetstvuyushchih fshch. edinic v fn. yachejkah svoih podsistem, chto sohranyaet ee strukturu
i chem obespechivaetsya sposobnost' kletki vypolnyat' algoritmy fn. yacheek sistem bolee
vysokogo poryadka, kuda ona vhodit v kachestve fshch. makroedinicy. Vse reakcii biosinteza
(reakcii assimilyacii) idut soglasno obshchej teorii sistem s pogloshcheniem energii dvizheniya
v prostranstve, kotoraya, kak by uvyazaya v strukture sistemy kletki, preobrazuetsya v
energiyu svyazi mezhdu ee fn. yachejkami.
Drugoj tip reakcij - reakcii rasshchepleniya - proishodit s
odnovremennym umen'sheniem energii svyazi, preobrazuyushchejsya v energiyu dvizheniya. Pri
reakciyah dissimilyacii fshch. edinicy podsistem kletki, yavlyayushchiesya sistemnymi obrazovaniyami
bolee nizkogo poryadka, otfunkcionirovav, raspadayutsya na fshch. edinicy svoego podurovnya,
gotovye pri neobhodimosti vstupit' v novye sinteziruyushchie reakcii s cel'yu formirovaniya
novyh struktur - fshch. edinic bolee vysokogo organizacionnogo urovnya. Oba tipa reakcij
tesno vzaimosvyazany i predstavlyayut soboj edinyj process, napravlennyj na zapolnenie
fn. yacheek struktury kletki deesposobnymi sootvetstvuyushchimi fshch. edinicami, chto v itoge
obespechivaet podderzhanie na dolzhnom urovne fn. svojstv kletki, kak edinogo celogo.
Odnim iz osnovnyh i slozhnejshih tipov sinteziruyushchih reakcij
yavlyaetsya biosintez belkov, protekayushchij v kletke postoyanno v techenie vsego perioda
vremeni ee sushchestvovaniya. V processe funkcionirovaniya kletki chast' ee belkov, prinyavshih
uchastie v kataliticheskih reakciyah, postepenno denaturiruetsya, ih struktura, a
sledovatel'no i funkcii, narushayutsya i oni udalyayutsya iz svoih fn. yacheek, a zatem i iz
samoj kletki. Ih mesto v fn. yachejkah zanimayut vnov' sintezirovannye belkovye molekuly,
po svoim fn. svojstvam polnost'yu identichnye osvobodivshim im mesto fshch. edinicam. Uchityvaya,
chto sushchestvuet velikoe mnozhestvo tipov belkovyh molekul, mehanizm ih sintezirovaniya,
sovershenstvovavshijsya v techenie dolgogo vremeni, v konechnom itoge prevratilsya v
specializirovannuyu podsistemu kletki s chetkim perechnem algoritmov funkcionirovaniya.
Programma sinteza belkov, to est' informaciya ob ih strukture,
zapisannaya i hranyashchayasya v DNK, napravlyaetsya v ribosomy s pomoshch'yu informacionnyh RNK
(i-RNK), sinteziruyushchihsya na DNK i tochno kopiruyushchih ee strukturu. Kazhdoj aminokislote
sootvetstvuet uchastok cepi DNK iz treh ryadom stoyashchih nukleotidov: A-C-A (cistein),
T-T-T (lizin), A-A-C (lejcin) i t.d. CHislo vozmozhnyh kombinacij iz 4-h nukleotidov
po 3 ravno 64, hotya ispol'zuemyh aminokislot vsego 20. Posledovatel'nost' nukleotidov
i-RNK v tochnosti povtoryaet posledovatel'nost' nukleotidov odnoj iz cepej genozapisi,
pri etom s kazhdogo gena mozhno snyat' lyuboe chislo kopij RNK.
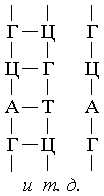 Perepisyvanie informacii na i-RNK, to est' process "transkripcii", proishodit
pri odnovremennom sintezirovanii i-RNK, kotoroe osushchestvlyaetsya s pomoshch'yu principa
komplementarnosti. V rezul'tate obrazuyushchayasya cepochka i-RNK po sostavu i posledovatel'nosti
svoih nukleotidov predstavlyaet soboj tochnuyu kopiyu sostava i posledovatel'nosti nukleotidov
odnoj iz cepej DNK. Molekuly i-RNK napravlyayutsya zatem k ribosomam, kuda postupayut takzhe
i aminokisloty, dostavlyaemye izvne kletki v uzhe gotovom vide. Aminokisloty popadayut
na ribosomu v soprovozhdenii transportnyh RNK (t-RNK), sostoyashchih v srednem iz 70 - 80
nukleotidnyh zven'ev, v 4 - 7 mestah komplementarnyh drug drugu. K odnomu iz koncov
t-RNK prisoedinyaetsya aminokislota, a v verhushke izgiba nahoditsya triplet nukleotidov,
kotoryj sootvetstvuet po kodu dannoj aminokislote. Na kazhduyu aminokislotu imeetsya svoya
t-RNK, to est' ih takzhe 20 raznovidnostej.
Perepisyvanie informacii na i-RNK, to est' process "transkripcii", proishodit
pri odnovremennom sintezirovanii i-RNK, kotoroe osushchestvlyaetsya s pomoshch'yu principa
komplementarnosti. V rezul'tate obrazuyushchayasya cepochka i-RNK po sostavu i posledovatel'nosti
svoih nukleotidov predstavlyaet soboj tochnuyu kopiyu sostava i posledovatel'nosti nukleotidov
odnoj iz cepej DNK. Molekuly i-RNK napravlyayutsya zatem k ribosomam, kuda postupayut takzhe
i aminokisloty, dostavlyaemye izvne kletki v uzhe gotovom vide. Aminokisloty popadayut
na ribosomu v soprovozhdenii transportnyh RNK (t-RNK), sostoyashchih v srednem iz 70 - 80
nukleotidnyh zven'ev, v 4 - 7 mestah komplementarnyh drug drugu. K odnomu iz koncov
t-RNK prisoedinyaetsya aminokislota, a v verhushke izgiba nahoditsya triplet nukleotidov,
kotoryj sootvetstvuet po kodu dannoj aminokislote. Na kazhduyu aminokislotu imeetsya svoya
t-RNK, to est' ih takzhe 20 raznovidnostej.
Sintez belkov i nukleinovyh kislot proishodit na osnove reakcij
matrichnogo sinteza. |tim obespechivaetsya pridanie vnov' obrazovannym soedineniyam fn.
svojstv zamenyaemyh fshch. edinic. Novye molekuly sinteziruyutsya v tochnom sootvetstvii
s planom, zalozhennym v strukture uzhe sushchestvuyushchih molekul. Poetomu v etih reakciyah
obespechivaetsya tochnaya, strogo specifichnaya posledovatel'nost' monomernyh zven'ev
v sinteziruemyh polimerah. Zdes' proishodit napravlennoe styagivanie monomerov v
opredelennoe mesto kletki - v fn. yachejki vnov' obrazuemogo polimera, pri etom
raspolozhenie samih fn. yacheek predopredelyaetsya strukturnoj organizaciej kopiruemoj
matricy. Rol' matricy v matrichnyh reakciyah igrayut makromolekuly nukleinovyh kislot DNK
i RNK. Monomernye molekuly (nukleotidy ili aminokisloty) v sootvetstvii s principom
komplementarnosti raspolagayutsya i fiksiruyutsya na matrice v strogo opredelennom, zadannom
poryadke. Zatem proishodit "sshivanie" monomernyh zven'ev v polimernuyu cep',
i gotovyj polimer sbrasyvaetsya s matricy. Posle etogo matrica gotova k sborke novoj
polimernoj molekuly. Pri pomoshchi matrichnogo tipa reakcij osushchestvlyaetsya vosproizvedenie
odnotipnyh soedinenij - fshch. edinic dannoj sistemy. Potrebnost' vosproizvedeniya odnotipnyh
fshch. edinic proslezhivaetsya na vseh urovnyah organizacii Materii i yavlyaetsya odnoj iz glavnyh
zakonomernostej obshchej teorii sistem.
Informaciya o strukture belka, zapisannaya v i-RNK v vide
posledovatel'nosti nukleotidov, perenositsya dalee v vide posledovatel'nosti aminokislot
v sinteziruemuyu polipeptidnuyu cep', to est' proishodit process "translyacii".
Po mere sborki belkovoj molekuly ribosoma polzet po i-RNK, za nej vtoraya, tret'ya i t.d.
Kazhdaya iz nih sinteziruet odin i tot zhe belok, zaprogrammirovannyj na dannoj i-RNK.
Kogda ribosoma projdet i-RNK ot konca do konca - sintez belka okonchen. Posle etogo
ribosoma idet na druguyu i-RNK, a belok po endoplazmaticheskoj seti napravlyaetsya
v sootvetstvuyushchuyu ego fn. svojstvam svobodnuyu fn. yachejku, kotoruyu on zapolnyaet
v kachestve fshch. edinicy.
Sintez belkov v kletke proishodit nepreryvno. Vse ribosomy,
pomeshchayushchiesya odnovremenno na odnoj i-RNK, ob®edinyayutsya v poliribosomu. Ribosoma rabotaet
vdol' i-RNK "shazhkami": triplet za tripletom RNK nahoditsya v kontakte s neyu. Dlya
sshivaniya polipeptidnoj cepi v ribosome imeetsya belok - sintetaza. Molekuly t-RNK, prohodya
cherez ribosomu, zadevayut svoim kodovym koncom mesto kontakta ribosomy s i-RNK. Esli
kodovyj triplet t-RNK okazhetsya komplementarnym k tripletu i-RNK, aminokislota,
dostavlennaya t-RNK, perehodit iz ee fn. yachejki v fn. yachejku sinteziruemoj molekuly
belka, takim obrazom stanovyas' fshch. edinicej ee struktury. |tim obespechivaetsya naivazhnejshee
pravilo postroeniya fn. sistem - pomeshchenie dannoj fshch. edinicy v strogo sootvetstvuyushchuyu ej
fn. yachejku ili, naoborot, zapolnenie fn. yachejki strogo sootvetstuyushchej ej fshch. edinicej.
Poetomu imeyushchijsya v lyuboj kletke mehanizm sintezirovaniya belkov obespechivaet stoprocentnuyu
garantiyu togo, chto perenosimaya t-RNK dannaya aminokislota popadet tol'ko v sootvetstvuyushchuyu
ej fn. yachejku struktury belka ili, naoborot, togo, chto v podoshedshuyu v ribosome
nezapolnennuyu ocherednuyu fn. yachejku sinteziruemogo belka popadet tol'ko
sootvetstvuyushchaya ej po fn. svojstvam fshch. edinica - trebuemaya aminokislota.
Zapolniv ocherednuyu fn. yachejku sinteziruemogo belka, ribosoma
delaet eshche odin shag po i-RNK, poluchaya takim obrazom informaciyu o fn. svojstvah sleduyushchej
fn. yachejki zapolnyaemoj struktury. t-RNK s osvobodivshejsya rabochej t-fn. yachejkoj uhodit
vo vnutrikletochnoe prostranstvo, gde zahvatyvaet sootvetstvuyushchuyu ej novuyu molekulu
aminokisloty s tem, chtoby vnov' nesti ee k lyuboj iz fshch. ribosom.
Molekuly belkov sinteziruyutsya v srednem okolo 1 - 2 min.
Process etot protekaet v prodolzhenie vsego perioda sushchestvovaniya kletki. Vse reakcii
belkovogo sinteza kataliziruyutsya special'nymi fermentami, vplot' do reakcij zahvata
t-RNK. Vse reakcii sinteza endotermichny, i poetomu kazhdoe zveno biosinteza vsegda
sopryazheno s rashodom ATF.
Lyubaya kletka sohranyaet svoj sostav i vse svoi fn. svojstva
na otnositel'no postoyannom urovne. Tak, soderzhanie ATF v kletkah sostavlyaet 0,04% i eta
velichina stojko uderzhivaetsya. Vklyuchenie i vyklyuchenie processov, obespechivayushchih podderzhanie
fn. svojstv kletki, proishodit v nej avtomaticheski. Osnovoj avtoregulyacii etih processov
yavlyaetsya special'naya signal'naya podsistema kletki, ispol'zuyushchaya s etoj cel'yu fn. svojstva
fshch. edinic predydushchih podurovnej, to est' elektromagnitnye svojstva elektronov, atomov
i t.d. Poetomu v lyuboj kletke vsegda imeetsya opredelennoe kolichestvo razlichnyh ionov i
drugih zaryazhennyh chastic, kotorye v celom sozdayut bioelektricheskie cepi, mikropolya i t.p.
Izmenenie bioelektricheskogo potenciala hotya by v odnom iz zven'ev lyuboj podsistemy kletki
sluzhit signalom dlya nachala ili okonchaniya toj ili inoj biohimicheskoj reakcii, togo ili
inogo peremeshcheniya fshch. edinic po fn. yachejkam razlichnyh podsistem kletki. Nalichie v
strukture kletok podsistemy signal'noj bioelektricheskoj svyazi, a takzhe ispol'zovanie
v etih celyah fn. svojstv fshch. edinic nizhnih podurovnej (elektronov, ionov i dr.) sluzhit
eshche odnim podtverzhdeniem nalichiya tesnoj vzaimsvyazi vseh urovnej edinoj sistemnoj
organizacii razvivayushchejsya Materii.
Itak, konechnym itogom Razvitiya Materii po urovnyu Z yavilos'
obrazovanie slozhnejshej sistemnoj struktury - organicheskoj kletki. Struktura kazhdoj kletki
vklyuchaet v sebya strogo opredelennoe kolichestvo razlichnyh fn. podsistem, kazhdaya iz kotoryh
vypolnyaet strogo opredelennuyu, prisushchuyu tol'ko ej funkciyu, obespechivayushchuyu normal'noe
funkcionirovanie vsej kletki v celom. Kazhdaya podsistema kletki imeet svoyu, strogo
opredelennuyu strukturu, vklyuchayushchuyu sistemnye obrazovaniya bolee nizkogo organizacionnogo
urovnya, imeyushchie polimolekulyarnoe stroenie so svoimi specificheskimi zakonami
funkcionirovaniya. Kazhdaya molekulyarnaya struktura vklyuchaet v sebya atomnye sistemy so
svoimi specificheskimi zakonami funkcionirovaniya. Atomnye struktury osnovany na zakonah
funkcionirovaniya subatomnyh podsistem. I tak beskonechno v strukturnuyu glub' Materii.
Vse ukazannoe nagromozhdenie fn. sistem i podsistem tonchajshim obrazom organizovano v
prostranstve i vo vremeni s odnoj edinstvennoj cel'yu - obespechit' proyavlenie v strogo
opredelennom meste v strogo opredelennyj period vremeni fn. svojstv svoeobraznogo
material'nogo obrazovaniya - organicheskoj kletki.
S etogo momenta Materiya vstupila v novyj etap svoego
kachestvennogo razvitiya - sozdanie samoreguliruyushchihsya i samoupravlyaemyh makrobiosistem.

[ Oglavlenie ]
[ Prodolzhenie teksta ]