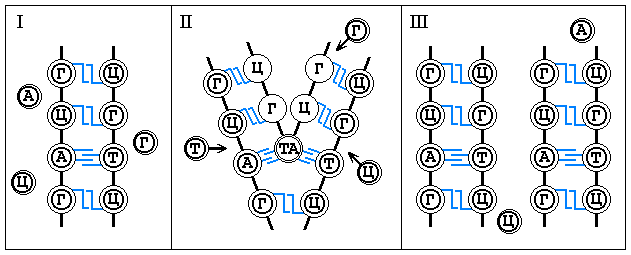
Благодаря принципу комплементарности, лежащему в основе
построения двойной спирали ДНК, молекула ДНК способна к удвоению. Во время этого
процесса две цепи разъединяются, образуя при этом две двойные цепи фн. ячеек, у которых
лишь один ряд заполнен фщ. единицами, а другой становится свободным. На следующем этапе
неассоциированные нуклеотиды из околосистемной среды заполняют соответствующие им
свободные фн. ячейки в обеих спиралях. В результате редупликации вместо одной молекулы
ДНК возникает две такого же точно нуклеотидного состава, как и первоначальная. Одна цепь
в каждой вновь образовавшейся молекуле ДНК остается от первоначальной молекулы, другая
синтезируется вновь. Таким путем вместе со структурой происходит передача фн. свойств
ДНК от материнской клетки к дочерней.
Графически это выглядит следующим образом:
Молекулы РНК также являются полимерами, как и ДНК,
но в отличие от них имеют одну спираль фн. ячеек, а не две. РНК выполняет в клетке
ряд функций, в том числе
1) транспортную (транспортируют
аминокислотные блоки к месту синтеза белков);
2) информационную (переносят
информацию о структуре белка);
3) рибосомную.
Еще одним очень важным нуклеотидом в структуре живой клетки
является аденозинтрифосфорная кислота - АТФ, содержание которой в клетках колеблется
от 0,04 до 0,2 - 0,5%. Его особенность состоит в том, что при отщеплении одной молекулы
фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорную кислоту) с выделением 40 кдж
энергии с 1 гр-молекулы.
Все указанные выше органические вещества являются сложными по
своей структуре и системной организации образованиями, но и они, в свою очередь, входят
в качестве фщ. единиц в фн. подсистемы интегрированной системы клетки. К числу основных
подсистем клетки отосятся следующие:
Наружная мембрана клетки. Регулирует поступление
ионов и молекул в структуру клетки и выход их из нее в околосистемную среду. Такой обмен
молекулами и ионами, то есть различными фщ. единицами, между системой клетки и внешней
средой происходит постоянно. Различают фагоцитоз - поглощение мембраной крупных частичек
вещества и пиноцитоз - поглощение воды и водного раствора. Через наружную мембрану из
клетки выводятся продукты ее жизнедеятельности, то есть отфункционировавшие в подсистемах
клетки фщ. единицы.
Цитоплазма. Внутренняя полужидкая среда клетки,
в системном объеме которой развернута внутренняя структура клетки, то есть ее ядро, все
органоиды (или органеллы), включения и вакуоли. Цитоплазма состоит из воды с растворенными
солями и разнообразными органическими веществами, среди которых преобладают белки.
Структура цитоплазмы состоит из фн. ячеек, не связанных жестко и свободно перемещающихся
во всем ее объеме. Заполняющие их фщ. единицы, когда это необходимо, переходят из них в
фн. ячейки органоидов. Поэтому основными функциями цитоплазмы являются аккумулятивная и
транспортная.
Эндоплазматическая сеть. Органоид клетки,
представляющий собой сложную систему каналов и полостей, пронизывающих всю цитоплазму
клетки. На мембранах гладкой эндоплазматической сети происходит синтез жиров и углеводов,
которые накапливаются в аккумулятивных фн. ячейках ее каналов и полостей, а затем
транспортируются к различным органоидам клетки, где они в качестве фщ. единиц занимают
соответствующие фн. ячейки их структур. На мембранах каналов и полостей располагается
также множество мелких округлых телец-рибосом.
Каждая рибосома состоит из двух мелких частиц, в состав которых
входят белки и РНК. В каждой клетке содержится по нескольку тысяч рибосом. На рибосомах
синтезируются все белки, входящие в состав данной клетки, путем сборки белковых молекул
из аминокислот, имеющихся в цитоплазме. Синтез белков - это сложный процесс заполнения
аминокислотными блоками соответствующих фн. ячеек их структур, который осуществляется
одновременно группой из нескольких десятков рибосом, или полирибосомой. Синтезированные
белки сначала накапливаются в каналах и полостях гранулярной эндоплазматической сети, а
затем транспортируются к тем подсистемам клетки, где расположены предназначенные для них
фн. ячейки. Эндоплазматическая сеть и полирибосомы представляют собой единый механизм
биосинтеза, аккумулирования и транспортировки белков.
Митохондрии. Органоид, основная функция
которого состоит в синтезе АТФ, представляющей универсальный источник энергии, необходимой
для осуществления постянно протекающих внутри клетки химических процессов. Количество
митохондрий в клетке колеблется от нескольких до сотен тысяч. Внутри митохондрий находятся
рибосомы и нуклеиновые кислоты, а также большое количество разнообразных ферментов.
Синтезированная АТФ заполняет транспортные фн. ячейки цитоплазмы и направляется к ядру
и органоидам клетки.
Пластиды. Органоиды растительных клеток.
Бывают различных типов. С помощью одного из них - хлоропластов, благодаря входящему
в их состав пигменту (хролофиллу), клетки растений способны, используя световую энергию
Солнца, синтезировать из неорганических веществ органические (углеводы). Этот процесс,
как известно, носит название фотосинтеза.
Комплекс Гольджи. Органоид всех растительных
и животных клеток, в котором происходит накопление белков, жиров и углеводов с последующей
их транспортировкой в соответствующие фн. ячейки как внутри клетки, так и вне ее.
Лизосомы. Органоид, имеющийся во всех клетках
и состоящий из комплекса ферментов, способных расщеплять белки, жиры и углеводы. В этом
заключается основная функция лизосом. В каждой клетке сосредоточены десятки лизосом,
участвующих в расщеплении уже отфункционировавших или аккумулятивных системных
образований, а также тех, которые попадают в клетку извне путем фагоцитоза и пиноцитоза.
В результате расщепления фщ. единицы покидают фн. ячейки расщепляемых структур, собираются
в фн. ячейках аккумулятивных систем данной клетки, а затем транспортируются в фн. ячейки
ее новых системных образований. Расщепленные с помощью лизосом отфункционировавшие
структуры клетки удаляются за ее пределы. Образование новых лизосом происходит в клетке
постоянно. Ферменты, функционирующие в лизосомах, как и всякие другие белки, синтезируются
на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети
к комплексу Гольджи, в полостях и трубочках которого формируются фн. ячейки структур
лизосом. Сформировавшись, лизосомы отделяются от концов трубочек и поступают в цитоплазму.
Клеточный центр. Органоид, расположенный в
одном из участков уплотненной цитоплазмы. В него входят две центриоли, играющие важную
роль при делении клетки.
Структура клеток содержит и другие органоиды: жгутики,
реснички и т.п., а также клеточные включения (углеводы, жиры, белки).
Вместе с тем, клетки, будучи сами по себе очень сложными
системными образованиями, в свою очередь являются фщ. единицами, заполняющими фн.
ячейки гиперсистем последующих уровней организации Материи. Вследствие этого в
системной организации клеток предусмотрен механизм, позволяющий за сравнительно
короткий период времени создавать аналогичные им системные формирования. В результате
клеточный цикл включает два периода:
1) Деление (митоз), в процессе которого
образуются две дочерние клетки;
2) Период между двумя делениями - интерфаза -
собственно время функционирования клетки.
Большую роль в делении клетки играет ее ядро, имеющееся
в каждой клетке и представляющее собой сложную фн. подсистему. Ядро имеет ядерную
оболочку, через которую в него и из него поступают белки, углеводы, жиры, нуклеиновые
кислоты, вода и разнообразные ионы. Попав внутрь ядра, они заполяют фн. ячейки ядерного
сока, а также ядрышек и хроматина. В ядрышках происходит синтез РНК, сами же они
формируются только в интерфазе. Хроматин представляет собой однородное вещество,
служащее аккумулятивной подсистемой, с помощью которой осуществляется формирование
хромосом при делении ядер.
Хромосомы являются основным механизмом клетки, где
собирается, хранится и выдается так называемая наследственная информация, включающая
в себя химическую запись последовательности фн. ячеек в структурах белков данной клетки.
Указанная информация хранится в находящихся в хромосомах молекулах ДНК. Таким образом,
молекулы ДНК представляют собой химическую запись структур всего разнообразия белков.
На длинной нити молекулы ДНК одна за другой следует запись информации о
последовательности фн. ячеек структур разных белков. Отрезок ДНК, содержащий информацию
о структуре одного белка, принято называть геном. Молекула ДНК представляет собой
собрание нескольких сот или тысяч генов. Диаметр хромосом невелик и составляет в среднем
140 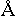 , длина же
их, повторяя длину молекул ДНК, может быть свыше 1 мм. В середине периода интерфазы
происходит синтез ДНК, в результате которого хромосома удваивается.
, длина же
их, повторяя длину молекул ДНК, может быть свыше 1 мм. В середине периода интерфазы
происходит синтез ДНК, в результате которого хромосома удваивается.
Важнейшая функция хромосом - быть хранительницей записей
структур и, соответственно, алгоритмических способностей фн. подсистем клетки с помощью
механизма образования белковых фщ. единиц. С течением времени, по мере приращения функций
того или иного вида органических систем, изменяется и совершенствуется запись в хромосомах,
следуя требованиям законов фн. развития Материи. В прямой зависимости от молекулярной
записи ДНК хромосом через механизм синтезирования белковых молекул происходят все
процессы жизнедеятельности клеток. Число хромосом постоянно для каждого вида животных
и растений, то есть в каждой клетке любого организма, принадлежащего к одному виду,
содержится совершенно определенное число хромосом (рожь - 14, человек - 46, курица -
78 и т.д.). Хромосомный набор, содержащийся в ядре одной клетки, имеет всегда парные
хромосомы. Так, 46 хромосом человека образуют 23 пары, в каждой из которых объединены
две одинаковые хромосомы. Хромосомы разных пар отличаются друг от друга по форме и месту
расположения. В результате митоза получаются две дочерние клетки, по строению полностью
сходные с материнской. Каждая из них имеет точно такие же хромосомы и такое же их число,
как и материнская клетка. Таким путем обеспечивается полная передача всей наследственной
информации каждому из дочерних ядер. Ядро и все органоиды цитоплазмы клетки
взаимодействуют как единая система.
Все клетки имеют сходный тип строения: ядро, митохондрии,
комплекс Гольджи, эндоплазматическую сеть, рибосомы и другие органоиды. Однако, прежде
чем стать столь совершенной системой, какой она является в наши дни, клетка прошла
длительный путь эволюции, отмеченный соответствующими отрезками на ординатах t и
ft тензора Развития Материи. Вначале она была частью неизвестных нам
неклеточных организмов, затем несовершенных одно- и многоклеточных, включая бактерии
и синезеленые водоросли, и в конечном итоге достигла совершенства сложного клеточного
механизма, характерного для представителей современного нам растительного и животного
мира. Вследствие движения Материи по ординате качества в процессе эволюции клетки
образовалось огромное разнообразие ее типов, каждый из которых был наделен строго
определенными фн. свойствами и соответствует определенной точке на данной ординате.
Вместе с тем, с определенного момента этот процесс стал
протекать одновременно с началом развития фн. систем более высокого организационного
уровня, фн. ячейки которых клетки стали заполнять в качестве фщ. единиц. В итоге клетка
превратилась в сложное системное образование, для поддержания фн. свойств которого внутри
и вне ее постоянно протекают сложные химические процессы. Постоянство процессов связано с
тем, что время функционирования фщ. единиц по мере увеличения их молекулярного веса все
более не совпадает со временем существования фн. ячеек структур, которые они заполняют,
так как при ограниченном пространстве перемещения фщ. единиц время их существования
находится в прямой зависимости от их фщ. массы. Кроме того, постоянство процессов вызвано
тем, что большинство протекающих в клетке химических реакций имеет необратимый характер.
Для всех этих реакций свойственны величайшая организованность и упорядоченность: каждая
реакция протекает в строго определенном месте в строго определенное время в строго
определенной последовательности. Молекулы ферментов расположены на мембранах митохондрий
и эндоплазматической сети в том порядке, в котором идут реакции.
В клетке имеются около тысячи ферментов, с помощью которых
протекают два типа реакций: синтеза и расщепления. Основным (созидающим)
типом реакций следует считать реакции синтеза, в процессе которых образуются сложные
молекулярные соединения, заполняющие в качестве фщ. единиц фн. ячейки подсистемных
структур клетки. Так, на замену каждой отфункционировавшей молекулы белка, покинувшей
ту или иную фн. ячейку, на освободившееся место поступает новая молекула белка, по
структуре и химическому составу, а следовательно и своим фн. свойствам, полностью
адекватная предыдущей фщ. единице. Это означает, что вновь синтезированная фщ. единица
способна (или должна быть способна) принять полноценное участие в любом из характерных
для данной фн. ячейки алгоритмов.
Синтезирование фщ. единиц осуществляется с помощью
функционирования специальных подсистем клетки на основе кодированной генозаписи ДНК.
Имеющие при этом место флуктуационные отклонения в случае их положительного эффекта
обратной связью фиксируются в генозаписи и служат целям дальнейшего совершенствования
данной системной структуры. При отрицательном эффекте от вновь синтезированной фщ.
единицы нарушается выполнение части фн. алгоритмов и, в случае, если система не в
состоянии это элиминировать, ненадлежащее функционирование соответствующей подсистемы
в конечном итоге может привести к разрушению структуры данной клетки в целом. Таким
путем организационное устройство клетки позволяет ей поддерживать постоянное наличие
соответствующих фщ. единиц в фн. ячейках своих подсистем, что сохраняет ее структуру
и чем обеспечивается способность клетки выполнять алгоритмы фн. ячеек систем более
высокого порядка, куда она входит в качестве фщ. макроединицы. Все реакции биосинтеза
(реакции ассимиляции) идут согласно общей теории систем с поглощением энергии движения
в пространстве, которая, как бы увязая в структуре системы клетки, преобразуется в
энергию связи между ее фн. ячейками.
Другой тип реакций - реакции расщепления - происходит с
одновременным уменьшением энергии связи, преобразующейся в энергию движения. При
реакциях диссимиляции фщ. единицы подсистем клетки, являющиеся системными образованиями
более низкого порядка, отфункционировав, распадаются на фщ. единицы своего подуровня,
готовые при необходимости вступить в новые синтезирующие реакции с целью формирования
новых структур - фщ. единиц более высокого организационного уровня. Оба типа реакций
тесно взаимосвязаны и представляют собой единый процесс, направленный на заполнение
фн. ячеек структуры клетки дееспособными соответствующими фщ. единицами, что в итоге
обеспечивает поддержание на должном уровне фн. свойств клетки, как единого целого.
Одним из основных и сложнейших типов синтезирующих реакций
является биосинтез белков, протекающий в клетке постоянно в течение всего периода
времени ее существования. В процессе функционирования клетки часть ее белков, принявших
участие в каталитических реакциях, постепенно денатурируется, их структура, а
следовательно и функции, нарушаются и они удаляются из своих фн. ячеек, а затем и из
самой клетки. Их место в фн. ячейках занимают вновь синтезированные белковые молекулы,
по своим фн. свойствам полностью идентичные освободившим им место фщ. единицам. Учитывая,
что существует великое множество типов белковых молекул, механизм их синтезирования,
совершенствовавшийся в течение долгого времени, в конечном итоге превратился в
специализированную подсистему клетки с четким перечнем алгоритмов функционирования.
Программа синтеза белков, то есть информация об их структуре,
записанная и хранящаяся в ДНК, направляется в рибосомы с помощью информационных РНК
(и-РНК), синтезирующихся на ДНК и точно копирующих ее структуру. Каждой аминокислоте
соответствует участок цепи ДНК из трех рядом стоящих нуклеотидов: А-Ц-А (цистеин),
Т-Т-Т (лизин), А-А-Ц (лейцин) и т.д. Число возможных комбинаций из 4-х нуклеотидов
по 3 равно 64, хотя используемых аминокислот всего 20. Последовательность нуклеотидов
и-РНК в точности повторяет последовательность нуклеотидов одной из цепей генозаписи,
при этом с каждого гена можно снять любое число копий РНК.
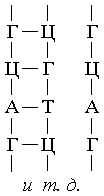 Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.
Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.
Синтез белков и нуклеиновых кислот происходит на основе реакций
матричного синтеза. Этим обеспечивается придание вновь образованным соединениям фн.
свойств заменяемых фщ. единиц. Новые молекулы синтезируются в точном соответствии
с планом, заложенным в структуре уже существующих молекул. Поэтому в этих реакциях
обеспечивается точная, строго специфичная последовательность мономерных звеньев
в синтезируемых полимерах. Здесь происходит направленное стягивание мономеров в
определенное место клетки - в фн. ячейки вновь образуемого полимера, при этом
расположение самих фн. ячеек предопределяется структурной организацией копируемой
матрицы. Роль матрицы в матричных реакциях играют макромолекулы нуклеиновых кислот ДНК
и РНК. Мономерные молекулы (нуклеотиды или аминокислоты) в соответствии с принципом
комплементарности располагаются и фиксируются на матрице в строго определенном, заданном
порядке. Затем происходит "сшивание" мономерных звеньев в полимерную цепь,
и готовый полимер сбрасывается с матрицы. После этого матрица готова к сборке новой
полимерной молекулы. При помощи матричного типа реакций осуществляется воспроизведение
однотипных соединений - фщ. единиц данной системы. Потребность воспроизведения однотипных
фщ. единиц прослеживается на всех уровнях организации Материи и является одной из главных
закономерностей общей теории систем.
Информация о структуре белка, записанная в и-РНК в виде
последовательности нуклеотидов, переносится далее в виде последовательности аминокислот
в синтезируемую полипептидную цепь, то есть происходит процесс "трансляции".
По мере сборки белковой молекулы рибосома ползет по и-РНК, за ней вторая, третья и т.д.
Каждая из них синтезирует один и тот же белок, запрограммированный на данной и-РНК.
Когда рибосома пройдет и-РНК от конца до конца - синтез белка окончен. После этого
рибосома идет на другую и-РНК, а белок по эндоплазматической сети направляется
в соответствующую его фн. свойствам свободную фн. ячейку, которую он заполняет
в качестве фщ. единицы.
Синтез белков в клетке происходит непрерывно. Все рибосомы,
помещающиеся одновременно на одной и-РНК, объединяются в полирибосому. Рибосома работает
вдоль и-РНК "шажками": триплет за триплетом РНК находится в контакте с нею. Для
сшивания полипептидной цепи в рибосоме имеется белок - синтетаза. Молекулы т-РНК, проходя
через рибосому, задевают своим кодовым концом место контакта рибосомы с и-РНК. Если
кодовый триплет т-РНК окажется комплементарным к триплету и-РНК, аминокислота,
доставленная т-РНК, переходит из ее фн. ячейки в фн. ячейку синтезируемой молекулы
белка, таким образом становясь фщ. единицей ее структуры. Этим обеспечивается наиважнейшее
правило построения фн. систем - помещение данной фщ. единицы в строго соответствующую ей
фн. ячейку или, наоборот, заполнение фн. ячейки строго соответстующей ей фщ. единицей.
Поэтому имеющийся в любой клетке механизм синтезирования белков обеспечивает стопроцентную
гарантию того, что переносимая т-РНК данная аминокислота попадет только в соответствующую
ей фн. ячейку структуры белка или, наоборот, того, что в подошедшую в рибосоме
незаполненную очередную фн. ячейку синтезируемого белка попадет только
соответствующая ей по фн. свойствам фщ. единица - требуемая аминокислота.
Заполнив очередную фн. ячейку синтезируемого белка, рибосома
делает еще один шаг по и-РНК, получая таким образом информацию о фн. свойствах следующей
фн. ячейки заполняемой структуры. т-РНК с освободившейся рабочей т-фн. ячейкой уходит
во внутриклеточное пространство, где захватывает соответствующую ей новую молекулу
аминокислоты с тем, чтобы вновь нести ее к любой из фщ. рибосом.
Молекулы белков синтезируются в среднем около 1 - 2 мин.
Процесс этот протекает в продолжение всего периода существования клетки. Все реакции
белкового синтеза катализируются специальными ферментами, вплоть до реакций захвата
т-РНК. Все реакции синтеза эндотермичны, и поэтому каждое звено биосинтеза всегда
сопряжено с расходом АТФ.
Любая клетка сохраняет свой состав и все свои фн. свойства
на относительно постоянном уровне. Так, содержание АТФ в клетках составляет 0,04% и эта
величина стойко удерживается. Включение и выключение процессов, обеспечивающих поддержание
фн. свойств клетки, происходит в ней автоматически. Основой авторегуляции этих процессов
является специальная сигнальная подсистема клетки, использующая с этой целью фн. свойства
фщ. единиц предыдущих подуровней, то есть электромагнитные свойства электронов, атомов
и т.д. Поэтому в любой клетке всегда имеется определенное количество различных ионов и
других заряженных частиц, которые в целом создают биоэлектрические цепи, микрополя и т.п.
Изменение биоэлектрического потенциала хотя бы в одном из звеньев любой подсистемы клетки
служит сигналом для начала или окончания той или иной биохимической реакции, того или
иного перемещения фщ. единиц по фн. ячейкам различных подсистем клетки. Наличие в
структуре клеток подсистемы сигнальной биоэлектрической связи, а также использование
в этих целях фн. свойств фщ. единиц нижних подуровней (электронов, ионов и др.) служит
еще одним подтверждением наличия тесной взаимсвязи всех уровней единой системной
организации развивающейся Материи.
Итак, конечным итогом Развития Материи по уровню З явилось
образование сложнейшей системной структуры - органической клетки. Структура каждой клетки
включает в себя строго определенное количество различных фн. подсистем, каждая из которых
выполняет строго определенную, присущую только ей функцию, обеспечивающую нормальное
функционирование всей клетки в целом. Каждая подсистема клетки имеет свою, строго
определенную структуру, включающую системные образования более низкого организационного
уровня, имеющие полимолекулярное строение со своими специфическими законами
функционирования. Каждая молекулярная структура включает в себя атомные системы со
своими специфическими законами функционирования. Атомные структуры основаны на законах
функционирования субатомных подсистем. И так бесконечно в структурную глубь Материи.
Все указанное нагромождение фн. систем и подсистем тончайшим образом организовано в
пространстве и во времени с одной единственной целью - обеспечить проявление в строго
определенном месте в строго определенный период времени фн. свойств своеобразного
материального образования - органической клетки.
С этого момента Материя вступила в новый этап своего
качественного развития - создание саморегулирующихся и самоуправляемых макробиосистем.

[ Оглавление ]
[ Продолжение текста ]